![[tmp928_thumb3.jpg]](http://lh3.ggpht.com/_X6JnoL0U4BY/S8HdkUtYtfI/AAAAAAAAYYI/fv2ArcWq3z0/s1600/tmp928_thumb3.jpg) |
| Line drawings of various larvae belonging to the suborder Myrmeleontiformia |
| Fossil and drawing of Parakseneura nigromacula's forewing |
 |
| Platystoechotes lineatus (Polystoechotidae), photographed in the Sierra Nevada by Beth Sands |
 |
| Oliarces clara, photographed by Shaun Winterton |
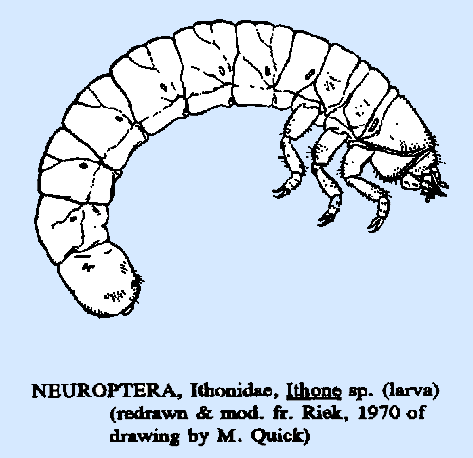 |
| Habitus of I. fusca grub |
 |
| Brown patches in a lawn—desecration! Blasphemy! Those irreligious Phyllophaga... |
 |
| A Californian P. punctata, photographed by Jim Moore |
Determining the solutions to this puzzle would be useful not just to neuropterists, but to anyone researching the United States' biotic history; and until we are so enlightened, our understanding of how our species has affected Earth's biosphere will be sadly incomplete.
*Sensory patches/bumps on insect wings, especially those of Neuroptera.
†Hairy, thickened portions of neuropteran wing margins.
‡Featuring mild temperatures with little variation.
________________________________________________________________
Aspöck, U.; Plant, J. D.; and Nemeschkal, H. L. (2001). Cladistic analysis of Neuroptera and their systematic position within Neuropterida (Insecta: Holometabola: Neuropterida: Neuroptera). Systematic Entomology, 26, 73-86.
Aspöck, U. (2002). Phylogeny of the Neuropterida (Insecta: Holometaboloa). Zoologica Scripta, 31, 51-55.
Aspöck, U. and Aspöck, H. (2008). Phylogenetic relevance of the genital sclerites of Neuropterida (Insecta: Holometabola). Systematic Entomology, 33, 97-127. Retrieved 5/13/13 from http://onlinelibrary.wiley.com/doi/10.1111/j.1365-3113.2007.00396.x/abstract
Carpenter, F. M. (1940). A Revision of the Neararctic Hemerobiidae, Berothidae, Sisyridae, Polystoechotidae, and Dilaridae (Neuroptera). Proceedings of the American Academy of Arts and Sciences, 74, 193-280.
Fabricius, J. (1793). Entomologica Systematica emendata et aucta secundum classes, ordines, genera, species, adjectis synonimis, locis observationibus, descriptionibus; tome #2. Hafniae: Christ. Gottl. Proft.
Faulkner, D. K. (1990). Current of the biology of the moth-lacewing Oliarces clara Banks (Insecta: Neuroptera: Ithonidae). Advances in Neuropterology. Proceedings of the Third National Symposium on Neuropterology, Pretoria, RSA; pp. 197-203.
Grebennikov, V. V. (2004). Grub-like larvae of Neuroptera (Insecta): a morphological review of the families Ithonidae and Polystoechotidae and a description of Oliarces clara. European Journal of Entomology, 101, 409-417. Retrieved 5/21/13 from http://www.eje.cz/pdfarticles/721/eje_101_3_409_IthonGreb.pdf
Gradstein, F. M.; Ogg, J. G.; and Smith, A. G. (2004). A Geologic Time Scale. Cambridge: Cambridge University Press.
Haring, E. and Aspöck, U. (2004). Phylogeny of the Neuropterida: a first molecular approach [electronic version]. Systematic Entomology, 29(3), 415-430. Retrieved 5/16/13 from http://onlinelibrary.wiley.com/doi/10.1111/j.0307-6970.2004.00263.x/abstract
Imms, A. D. (1957). A General Textbook of Entomology. London: Methuen.
Jepson, J. E.; Makarkin, V. N.; and Jarzembowski, E. A. (2009). New lacewings (Insecta: Neuroptera) from the Lower Cretaceous Wealden Supergroup of Southern England [electronic version]. Cretaceous Research, 30, 1325-1338. Retrieved 5/23/13 from http://www.academia.edu/1076267/New_lacewings_Insecta_Neuroptera_from_the_Lower_Cretaceous_Wealden_Supergroup_of_Southern_England
Johnson, R. (1992). Cheese-weed owlfly (Oliarces clara). Unpublished short abstract for United States Fish & Wildlife Service. Phoenix: Arizona Ecological Services.
Lambkin, K. J. (1988). A re-examination of Lithosmylidia Riek from the Triassic of Queensland with notes on Mesozoic "osmylid-like" fossil Neuroptera (Insecta: Neuroptera). Memoirs of the Queensland Museum, 25, 445-458.
MacLeod, E. G. (1964). A Comparative Morphological Study of the Head Capsule and Cervix of Larval Neuroptera (Insecta). Unpublished PhD thesis, Department of Biology, Harvard University, Cambridge, Massachusetts.
Makarkin, V. N. and Menon, F. (2005). New species of the Mesochrysopidae (Insecta, Neuroptera) from the Crato Formation of Brazil (Lower Cretaceous), with taxonomic treatments of the family [electronic version]. Cretaceous Research, 26(5), 801-812. Retrieved 5/10/13 from http://www.biosoil.ru/files/00000710.pdf
Makarkin, V. N.; Yang, Q.; and Ren, D. (2012). A new basal osmylid neuropteran insect from the Middle Jurassic of China linking Osmylidae to the Permian-Triassic Archeosmylidae [electronic version]. Acta Palaeontologica Polonica. Retrieved 5/23/13 from http://www.app.pan.pl/archive/published/app57/app20110018_acc.pdf
Makarkin, V. N. and Archibald, S. B. (2003). Family Affinity of the Genus Palaeopsychops Andersen with Description of a New Species from the Early Eocene of British Columbia, Canada (Neuroptera: Polystoechotidae). Annals of the Entomological Society of America, 96(3), 171-180. Retrieved 5/22/13 from http://www.brucearchibald.com/docs/Makarkin%20and%20Archibald%202003.pdf
Makarkin, V. N. and Archibald, S. B. (2009). A new genus and first Cenozoic fossil record of moth lacewings (Neuroptera: Ithonidae) from the Early Eocene of North America. Zootaxa, 2063, 55-63. Retrieved 5/12/13 from http://www.brucearchibald.com/docs/Makarkin_and_Archibald_2009.pdf
Marshall, S. (2006). Insects: Their Natural History and Diversity. Richmond Hill: Firefly Books.
Martynova, O. M. (1948). Mesozoic lacewings (Neuroptera) and their bearing on concepts of phylogeny and systematics of the order. Trudy Paleontologicheskogo Instituta Akademii Nauk SSSR, 7(1), 1-232.
Navás, L. (1930). Insectos neotropicos. Revista Chilena de Historia Natural, 33, 17-24.
Oswald, J. D. (1998). Rediscovery of Polystoechotes gazullai Navás (Neuroptera: Polystoechotidae). Proceeding of the Entomological Society of Washington, 100, 389-394.
Penny, N. D. (1996). A remarkable new genus and species of Ithonidae from Honduras (Neuroptera). Journal of Kansas Entomology, 69, 81-86.
Ponomarenko, A. G. (1992). Neuroptera (Insecta) from the Lower Cretaceous of Transbaikalia. Palaeontological Journal, 26(3), 56-66.
Ren, D. and Engel, M. S. (2008). Aetheogrammatidae, a new family of lacewings from the Mesozoic of China (Neuroptera: Myrmeleontiformia). Journal of the Kansas Entomological Society, 81(3), 161-167.
Riek, E. F. (1974). The Australian moth-lacewings (Neuroptera: Ithonidae). Journal of the Australian Entomological Society, 15, 37-54.
Tillyard, R. J. (1922). The life-history of the Australian moth-lacewing, Ithone fusca (Order Neuroptera, Planipennia). Bulletin of Entomological Research, 13, 205-223.
Welch, P. S. (1914). The early stages of the life history of Polystoechotes punctatus Fabr. Bulletin of the Brooklyn Entomological Society, 9, 1-6.
Winterton, S. L. and Makarkin, V. N. (2010). Phylogeny of Moth Lacewings and Giant Lacewings (Neuroptera: Ithonidae, Polystoechotidae) Using DNA Sequence Data, Morphology, and Fossils [electronic version]. Annals of the Entomological Society of America, 103(4), 511-522. Retrieved 5/11/13 from http://www.bioone.org/doi/abs/10.1603/AN10026
Yang, Q.; Makarkin, V. N.; Winterton, S. L.; Khramov, A. V.; and Ren, D. (2012). A Remarkable New Family of Jurassic Insects (Neuroptera) with Primitive Wing Venation and Its Phylogenetic Position in Neuropterida. PLOS One. Retrieved 5/9/13 from http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0044762
No comments:
Post a Comment