{kind=link}

Still, they have a degree of notoriety among entomologists, as was shown when I struck up a conversation with a fellow student at Ohio State University's Stone Laboratory some time ago, and happened to mention twisted-winged parasites.
"Yeah," my peer said, "those things are f*cking awesome."
 |
| A male member of Corioxenidae; photographed in Texas by Mike Quinn |
 |
| Female of Callipharixenos muiri (Kathirithamby, 1989) |
 |
| A male stylopid (lower left) humpin' an unseen female in a bee's butt; photographed by Colin Boyd |
 |
| A Tunisian female of Mengenilla moldrzyki (Mengenillidae); Pohl et al., 2012 |
 |
| A male Caenocholax fenyesi (Myrmecolacidae) eclosing from an ant (Solenopsis invicta); Cook et al., 2005 |
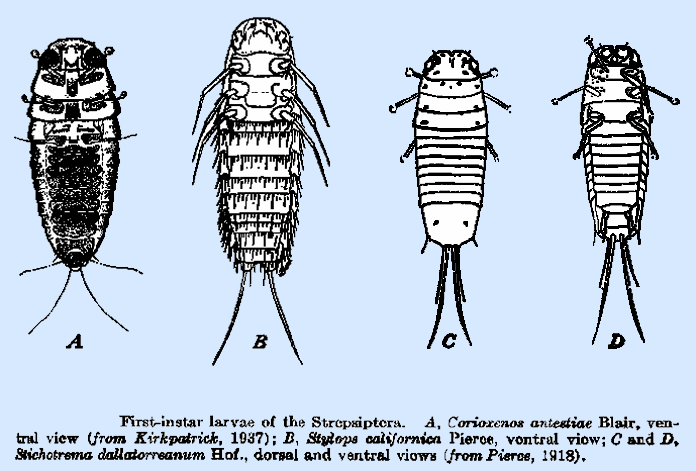
If you are familiar with the various insects that begin their lives as planidia, you will know that this initial ontogenetic period invariably corresponds with hypermetamorphosis (complete metamorphosis with subdivision of the larval stage). Hence you could correctly surmise that twisted-winged parasites are hypermetamorphic too (Osswald et al., 2010): once a planidium has penetrated its host (often following the host's molt—the exoskeleton is then more pliant) and plunged into the fluid-filled interior, subsequent instars take a sedentary foot-lacking grub-like shape; a protective bag of exuviae gradually surrounds the larva, since each time it molts it doesn't shed its cuticle. One species masquerades as a part of its katydid host by enclosing itself in a sack of tissue derived from the host's epidermis, thereby deflecting the katydid's immune reaction (Kathirithamby et al., 2003).
 |
| Stylops melittae pupae protruding from a Czech wasp; picture credit given to Josef Dvořák
|
 |
| Ripiphorus vierecki (Ripiphoridae), male, photographed by Margarethe Brummermann in Arizona |
But these characteristics do not really form a convincing argument for kinship between the two: many unrelated male insects have extravagantly feathery antennae; hypermetamorphosis has arisen independently in insects several times; and beetle elytra, however ineffectually stunted they happen to be in Ripiphoridae, are functionally dissimilar to the strepsipteran halteres (Pix et al., 1993). Still, the ad hoc placement stood until molecular phylogeny came along in the 1990s and introduced a rival hypothesis. Analyses using genetics (Chalwatzis et al., 1996; Whiting et al., 1997) indicated that the Strepsiptera were a sister-group to the true flies (Diptera), a view not without precedent (Newman, 1864; Pierce, 1918). Both taxa have a wing-pair modified into halteres—but the gyroscopic stabilizers originate from the metathorax in flies, as opposed to the mesothoracic ones in twisted-winged parasites: a big evolutionary jump, in developmental terms; but not in the realm of impossibility with the aid of homeotic mutation (Whiting & Wheeler, 1994).
The two allegedly comprised a clade dubbed "Halteria". This theory, while popular, was not without controversy; an argument was made that the grouping of Strepsiptera and Diptera together was an artifact due to a cladistic phenomenon called "long-branch attraction" (Carmean & Crespi, 1995; Huelsenbeck, 1997), which I will not deign to explain (since I really have no idea what it is): thus it seemed that Halteria was artificial, and strepsipterans' descent remained up in the air. A slew of new (and reputedly improved) morphological/molecular phylogenies (Beutel et al., 2010; McKenna & Farrell, 2010) have now strongly pointed towards twisted-winged parasites' being beetles' closest kin (although not beetles themselves, as some have persisted in suggesting) (Niehuis et al., 2012). For the time being, thus, we may say that beetles are twisted-winged parasites' nearest relatives.
*Emergence from the pupa.
†The internal bodily fluid that bathes the organs of arthropods, analogous to vertebrate blood and lymph.
‡ Larva-like but sexually mature.
____________________________________________________________
Beutel, R. G.; Friedrich, F.; Hörnschemeyer, T.; Pohl, H.; Hünefeld, F.; Beckmann, F.; Meier, R.; Misof, B.; Whiting, M. F.; and Vilhelmsen, L. Morphological and molecular evidence converge upon a robust phylogeny of the megadiverse Holometabola [electronic version]. Cladistics, 27(4), 341-355. Retrieved 2/24/13 from http://onlinelibrary.wiley.com/doi/10.1111/j.1096-0031.2010.00338.x/abstract
Bravo, F.; Pohl, H.; Silva-Neto, A.; and Beutel, R. G. (2009). Bahiaxenidae, a “living fossil” and a new family of Strepsiptera (Hexapoda) discovered in Brazil [electronic version]. Cladistics: the International Journal of the Willi Hennig Society, 25(6), 614-623.
Buschbeck, E.; Ehmer, B.; and Hoy, R. (1999). Chunk versus point sampling: visual imaging in a small insect. Science, 286, 1178-1180.
Carmean, C. and Crespi, B. J. (1995). Do long branches attract flies? Nature, 373, 666.
Crowson, R. A. (1960). The phylogeny of Coleoptera. Annual Review of Entomology, 5, 111-134.
Clausen, C. P. (1940). Entomophagous Insects. New York City: McGraw-Hill.
Chalwatzis, N.; Hauf, J.; van de Peer, Y.; Kinzelbach, R.; and Zimmermann, F. K. (1996). 18S ribosomal RNA genes of insects: primary structure of the genes and molecular phylogeny of the Holometabola. Annals of the Entomological Society of America, 89, 788-803.
Cook, J. L.; Calcaterra, L. and Nuñez, L. (2005). First record of Caenocholax fenyesi (Strepsiptera: Myrmecolacidae) parasitizing Solenopsis invicta (Hymenoptera: Formicidae) in Argentina, with a discussion of its distribution and host range. Entomological News, 115(2), 61.
Cook, J. L. and Derr, D. P. (2004). Antennal morphology of Caenocholax fenyesi (Strepsiptera: Myrmecolacidae) based on scanning electron microscopy. ESA Annual Meeting.
Drew, A. I. and Allwood, J. A. (1985). A new family of Strepsiptera parasitizing fruit flies (Tephritidae) in Australia. Systematic Entomology, 10, 129-134.
Esaki, T. and Miyamoto, S. (1965). The Strepsiptera parasitic on Heteroptera. Proceedings of the International Congress of Entomology, Montreal; 1, 375-381.
Huelsenbeck, J. P. (1997). Is the Felsenstein Zone a fly trap? Systematic Biology, 46, 69-74.
Johnston, J. S.; Ross, L. D.; Beani, L.; Hughes, D. P.; and Kathirithamby, J. (2004). Tiny genomes and endoreduplication in Strepsiptera. Insect Molecular Biology, 13(6), 581-585.
Kathirithamby, J. (1989). Review of the order Strepsiptera. Systematic Entomology, 14, 41-92.
Kathirithamby, J. (2000). Morphology of the female Myrmecolacidae (Strepsiptera) including the apron, and an associated structure analogous to the peritrophic matrix. Zoological Journal of the Linnean Society, 128, 269-287.
Kathirithamby, J. and Kifune, T. (1994). Strepsiptera (Insecta) parasitizing Onychostylus pallidiolus (Shiraki), the blattellid cockroach in southwestern-most Japan. Entomologist, 113, 217-219.
Kathirithamby, J.; McMahon, D. P.; Anober-Lantican, G. M.; and Ocampo, V. R. (2012). An unusual occurrence of multiparasitism by two genera of Strepsiptera (Insecta) in a mango leafhopper Idioscopus clypealis (Lethierry) (Hemiptera: Cicadellidae) in the Philippines [electronic version]. Zootaxa, 3268, 16-28. Retrieved 2/19/13 from http://mapress.com/zootaxa/2012/f/z03268p028f.pdf
Kathirithamby, J.; Ross, L. D.; and Johnston, J. S. (2003). Masquerading as Self? Endoparasitic Strepsiptera (Insecta) Enclose Themselves in Host-Derived Epidermal Bag. Proceedings of the National Academy of Sciences of the United States of America, 100(13), 7655-7659.
Kristensen, N. P. (1991). Phylogeny of extant hexapods. In Naumann, I. D.; Cornell, P. B. C.; Lawrence, J. F.; Neilson, E. S.; Spradberry, J. P.; Taylor, R. W.; Whitten, M. J.; and Littlejohn, M. J. (eds.): The Insects of Australia: a Textbook for Students and Research Workers (2nd ed.) (pp. 125-140). CSIRO, Melbourne: Melbourne University Press.
Maxumdar, A. and Chaudhuri, P. K. (1999). Strepsipteran insects of the genus Tridactylophagus Subramaniam from India (Strepsiptera: Halictophagidae). Journal of South Asian Natural History, 4(1), 13-17.
McKenna, D. D. and Farrell, B. D. (2010). 9-Genes Reinforce the Phylogeny of Holometabola and Yield Alternate Views on the Phylogenetic Placement of Strepsiptera. PLoS ONE, 5(7), e11887. Retrieved 2/24/13 from http://www.torna.do/s/9-genes-reinforce-the-phylogeny-of-holometabola-and-yield-alternate-views-on-the-phylogenetic-placement-of-Strepsiptera/
McQueen, R. (1998, June 1). Hitch-Hiking Insects [electronic version]. Creation. Retrieved 2/18/12 from http://www.answersingenesis.org/articles/cm/v20/n3/hitch-hiking-insects
Newman, E. (1864). Natural situation of Stylops among insects. Entomologist, 2, 231-232.
Niehuis, O.; Hartig, G.; Grath, S.; Pohl, H.; Lehmann, J.; Tafer, H.; Donath, A.; Krauss, V.; Eisenhardt, C.; Hertel, J.; Petersen, M.; Mayer, C.; Meusemann, K.; Peters, R. S.; Stadler, P. F.; Beutel, R. G.; Bornberg-Bauer, E.; McKenna, D. D.; and Misof, B. (2012). Genomic and Morphological Evidence Converge to Solve the Enigma of Strepsiptera. Current Biology, 22, 1-5.
Osswald, J.; Pohl, H.; and Beutel, R. G. (2010). Extremely miniaturized and highly complex: the thoracic morphology of the first instar larva of Mengenilla chobauti (Insecta, Strepsiptera). Arthropod Structure & Development, 39, 287-304.
Pierce, W. D. (1918). The comparative morphology of the order Strepsiptera. U.S. National Museum Proceedings, 54, 391-501.
Pix, W.; Nalbach, G.; and Zeil, J. (1993). Strepsipteran forewings are haltere-like organs of equilibrium. Naturwissenschaften, 80, 371-374.
Pohl, H. (2000). Die Primärlarvenächerflügler—evolutionäre Trends (Insecta, Strepsiptera). Kaupia, Darmstädter Beiträge zur Naturgeschichte, 10, 1-144.
Pohl, H. (2002). Phylogeny of the Strepsiptera based on morphological data of the first instar larvae. Zoologica Scripta, 31, 123-134.
Pohl, H. and Beutel, R. G. (2008). The evolution of Strepsiptera (Hexapoda). Zoology, 111(4), 318-338.
Pohl, H.; Niehuis, O.; Gloyna, K.; Misof, B.; and Beutel, R. G. (2012). A new species of Mengenilla (Insecta: Strepsiptera) from Tunisia. ZooKeys, 2012(198), 79-101. Retrieved 20/2/13 from http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3368257/
Rossi, P. (1793). Observation de M. Rossi sur un nouveaugenre d'Insecte, voisin des Ichneumons. Bulletin de la Société Philomatique à ses Correspondants, 1(49).
Silvestri, F. (1943). Studi sugli 'Strepsiptera' (Insecta). III. Descrizione e biologia di 6 specie italiane di Mengenilla. Boll. Lab. Zool. Gen. Agr. Fac. Agr. Portici, 32, 197–282.
Smith, G. and Hamm, A. H. (1914). Studies in the experimental analysis of sex, pt. II: On Stylops and stylopization. Quarterly Journal of Microscopic Science, 60, 435-461.
Subramanium, T. V. (1922). Some natural enemies of the mango leafhoppers (Idiocerus spp.) in India. Bull. Ent. Res., 12, 465-467.
Ulrich, W. (1930). Die Strepsipteren-Männchen als Insekten mit Halteren an Stelle der Vorderflügel. Zeitschrift für Morphologie und Ökologie der Tiere, 17, 552-624.
Whiting, M. F. (2003). Strepsiptera. In Resh, V. H. & Cardé, R. T. (eds.): Encyclopedia of Insects (pp. 1094-1096). Waltham: Academic Press.
Whiting, M. F. and Wheeler, W. C. (1994). Insect homeotic transformation. Nature, 368(696).
Whiting, M. F.; Carpenter, J. C.; Wheeler, Q. D.; and Wheeler, W. C. (1997). The Strepsiptera problem: phylogeny of the Holometabolous insect orders inferred from 18S and 28S ribosomal DNA sequences and morphology. Systematic Biology, 46, 1-68.
No comments:
Post a Comment